

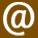


Abstract: This article uncovers the intricate relationships between biological evolution and life-acquired bodily characteristics arising from repetitive behaviors, calling into question previously dismissed theories of inheritance and providing a comprehensive overview of the deeper evolutionary forces that have shaped the diversity of genomes throughout the entire history of life. We will examine the role of repetitive behaviors, the force of gravity, and other hidden environmental forces which pressure life to move in ways that are involved in shaping the future evolution of living organisms. We will highlight a list of reasons why these ideas are proving to be more accurate than all other previous models of evolutionary theory. We will also conclude this disclosure with criticisms and controversies surrounding Lamarckian evolution, as well as the possibility of revisiting Jean-Baptiste de Lamarck’s work. Our research and experiments focus on identifying potential areas for future study, including epigenetic mechanisms and their implications for human health; deepening our understanding of the causes of inherited diseases; and providing new insights into evolutionary biology.
I. Introduction to the term “Biological Erosion”—One of the 5 Hidden Forces
Darwin’s theory of evolution is central to biology, but Jean-Baptiste Lamarck, a pioneer in the early 1800s, first proposed the idea that species change over deep time through physical transformations. His belief in gradual evolution and connected family relationships influenced scientific thought, though his mechanism of inheriting acquired traits was mocked. Recent advances in epigenetics have revived interest in Lamarck’s ideas, suggesting he may have been partially correct.
As we explore more of the complex relationships between biology, environment, and evolution, it’s essential to reconsider the potential validity of Lamarck’s theories, withholding any preconceived judgments, so that an unbiased new perspective can have a fair chance at being observed with its relevance to a modern understanding of the mysterious laws of epigenetics that are currently being detected and need answers.
One of the most consistent observations I keep seeing in all my work, throughout my life, when handling and observing animal anatomy, can be summarized with the word “biological erosion” (a new term I had to coin to verbally describe a phenomenon I keep spotting in every transitional genome’s bone morphology). If you are new to my work, I already briefly introduced a few examples of this observation without focusing so much on the 5 hidden forces that I am detailing here in part 2. If you are interested in reading my first introduction to this study, click this link to jump over to Part 1: What If We Created Ourselves, With The Motion of Our Own Bodies? [Part 1: Looking deeper into the world of epigenetics]
Hidden Force 1 —Biological Erosion / Entropy
Biological erosion is a term that describes areas in the skeleton that seem not only to have been shaped by the history of each living animal family’s movement in its chosen environment, but older features that took longer than one lifetime to take their current shape, indicating that these older features were inherited from the millions of preceding generations who were doing the same things to their bodies but in an earlier stage of the process. These include both the damages and the gains. These physical features, preserved through injury and damage, exhibit a remarkable resemblance to rocky landforms shaped by repetitive forces such as wind, rain, water currents, and features we normally associate with erosion. It reveals a direct connection to our daily habits, shaping the new future species we eventually become. These bodily adaptations flip on like a delayed switch over many generations after a genome had externally switched into a dramatically different diet, environment, or even simply just remained in its current environment but tried a brand-new behavior over an old one for another extension of time (i.e., tree-climbing squirrels to tree-glider squirrels).
You will see in this overview how consistent it is when you study the chronology of any vertebrate’s evolution, there will always be predictable signs of biological erosion in their bones, stamped and scraped by the wearing of time from the species’ full unanimous cooperative collective repetitious behaviors all working together as one machine hammering away and reshaping the bones into endless bodily forms.
The consistent appearance of fused or reinforced bones in species with habitual behaviors; such as the fused tarsals in runners, reinforced beak structures in peckers, or thickened limb bones in burrowers—all demonstrates a predictable pattern of biological erosion; where features are bent or carved into while the damage is healed by the body, which seems to come coupled together with synostosis (more on this later).
Microstructural analyses reveal that bones subjected to repetitive stress develop increased osteoblastic activity, thicker cortical bone, and reinforced trabeculae, all of which now appear to become part of the heritable genetic blueprint over time.
The presence of wear patterns, microcracks, and remodeling signs in fossils across diverse taxa supports the hypothesis that behavioral forces are encoded in skeletal morphology. We already know that wear and tear can occur in individuals as they age over a lifetime. That part is nothing new, but what is astonishing is finding that these same patterns persist into macro changes in the entire species’ future evolution. These phenomena are not isolated; they form a cohesive pattern across evolutionary history, reinforcing the idea that physical activity, environmental pressures, and habitual behaviors leave a biological legacy that influences future generations. What that means is that what you and your spouse were doing to your bodies and then passed down traditionally is being recorded in your DNA. What we are all doing to ourselves now, in the behaviour that shapes our collective current lives, will have a permanent effect on future human health, passed down through our offspring.
We will explore the evidence for this and define it more clearly for those who need to see it to believe it. Make no mistake about it: from land to sea and into the air, animals far and wide are all operating under this unbelievable uniform law that we have missed by ignoring and dismissing Lamarckian evolution. This marks a new era for epigenetic scientists, and the rapidly growing field of biophysics is inevitably going to reach even higher heights once this is accepted. Let us continue and explore the implications of these concepts for our understanding of evolution and adaptation.
My first article for Graham Hancock’s Article Forum, written under this same heading title, Part 1, only summarized my hypothesis and experimental process as a short introduction. Part 2 will now skim through all my notes and observations, which help open up a broader perspective on the details that consistently point to the same conclusion I am left with, and now have no choice but to declare as the truth of how a species’ bodies morph over time. This will not only revolutionize the field of genetics but also will sprout new branches for the field of biophysics.
What is Biological Erosion?
• Biological Erosion: is a force adjacent to the same force we observe and describe in geology when explaining the formation of a long river carved out by water erosion, melting glaciers sliding against terrain, mountain peaks rounding out, and rough desert sand blowing in the wind and grinding relentlessly against every land feature caught in its path. Biological erosion is the biological version of that same force within entropy. In a side-by-side comparison, it describes why so many biological and geological features look like veins or cracks branching out into fractals, and you will find it behind a host of explanations for the specific shapes and sizes of specialized bones in the body that undergo constant use by the driver of the body.
What is the study of biophysics?
• Biophysics: a small study, recognized as a branch in the field of biology, where the same laws in the world of physics, such as the laws of motion, chemistry, and thermodynamics, can be witnessed interacting in the biology of living specimens (with its potential capacity in tampering with the evolutionary process itself, as observed in this article). Biophysics recognizes entropy’s involvement in biomechanical and thermal systems, as well as ectropy when homeostasis is restored out of chaotic phases. It recognizes the role of gravity and chemistry in bone and cell structures. A biophysicist might pull data from a cooling dead carcass and observe cellular changes as the body temperature cools to room temperature after death, when it is no longer in a sustained system. Simply put, biophysics is an interdisciplinary science that combines principles from physics and biology to understand how biological systems work. It aims to research and explain the physical mechanisms underlying all processes in living organisms, such as how molecules move, how cells generate energy, and how muscles produce kinetic force to pull the weight of bones and produce movement and mobility. Darwin and Wallace’s natural and sexual selection are also among the external factors that act on genes when studying them through biophysics.
Selective pressures like these, which demand success without mercy, limit organisms to crafting bodies that benefit the long-term survival of the species. Hidden parameters like the laws for survival of the fittest (natural/sexual selection) are simply selective filters that interact with the evolutionary process by eliminating the older models of genes that do not fit anymore, due to the evolutionary mismatch of being in the wrong environment or having the wrong adaptations that do not complement a higher survival rate in the currently changed environment. Oftentimes, we see a direct relationship between the external environment switching on epigenetic expressions in an organism’s DNA, so it is safe to also conclude that epigenetics has already proven these hypotheses around acquired characteristics to be true, and this feature in our genes has a connected relationship with these environmental factors, as they both trigger each other’s responses. The environment needs the biosphere’s symbiosis to stay lush and tropical, and the biosphere needs to adapt to the environment, which is triggered via epigenetics to change along with it. Adaptation occurs over long periods of trial and error, but the ultimate outcome is biological and geological evolution of both genomes and their impact on environmental quality and symbiosis.
Examples of Biological Erosion:
In Part 1 of this article series I demonstrated only a few examples: the wingtip feathers in avian birds being carved out as if they were subject to heavy wind erosion, and the bone warping of saber-tooth animals like Thylacosmilus‘ double-edged chin, that coincides with the shape and length of its saber tooth canines; are both certainly great observations of biological erosion being recorded into the bones and soft tissues like feathers. The constant flapping and soaring in the wind, or the constant up-and-down rubbing between Thylacosmilus‘ canines and the sides of its chin, all caused the affected tissues’ shape to warp around and slide in the direction of the repetitive force exerted on it, as a hard clay would do under those same repetitive motions. The use of the wings or the mouth itself, with its overgrown feathers, or overgrown teeth attached, is why the wings were carved out by wind, or the smudged bony protrusions in the underside of Thylacosmilus‘ mandible began to get pushed around from the original shape of its earliest ancestor’s bodies.
To see how consistent this is, just look at every version of saber-tooth convergence in the history of all life on Earth. You will always see that, as the saber fangs reach a maximum girth, with every species that evolves them, the two sides of a low drooping chin will always show up after the girth of the tooth finally makes hard contact with the sides of its chin. These changes to the surface and mass of the bone follow a natural law that behaves exactly like erosion! Repetitive friction is what drives geological and biological erosion. The only difference between geological erosion and biological erosion is that the cells of bone and soft tissue are constantly self-healing as their surface erodes; while inanimate objects in geology such as limestone and granite do not heal and will continue to erode until nothing is left of its structure (i.e. a crack in a bone will heal into thick bone scarring versus a crack in a rock will only separate wider until it breaks into two). Biological erosion was the first hidden force I discovered, silently operating behind the curtains of genetic inheritance.
Hidden Force 2 —The External Laws of Physics / Epigenetics / Selective Pressures
Biophysics will be the future of understanding evolution more thoroughly. Its field will help us explore the structure and function of molecular exchanges in the body, the physical limits of cells, and the predictable changes to tissues as we age or evolve at a fundamental level. By incorporating concepts such as biological erosion and biophysics into our understanding of biomechanics/physiology, and charting all evolutionary changes, we can understand the roles that these physical, chemical, and energetic forces of nature play on the body’s living systems. When these forces affect the body’s systems, all the acquired changes get copied, and the motion of evolutionary change is set forward in each child born that mimics their parents’ behavior. Essentially, biophysics is about combining the tools and formulas of Newtonian physics with Rudolf Clausius’ laws of thermodynamics to better understand the mechanics driving the survival and evolution of life itself.
II. Biological Erosion and Synostosis: The Fusion & Slow Reshaping of Healing Bones
Hidden Force 3 —Synostosis & Returning to Homeostasis
As bones get damaged, from either breaks or stress from trauma caused by a heavier load or very different bodily movement, the damaged areas of the bone are reshaped by thick scarring or twists to their shape. Normally, we understand that these single-life injuries never get passed on to the individual’s children, but recent research in epigenetics and my own research suggest that some repetitive injuries actually do accumulate into massive changes within the skeletal morphology of the entire breeding population in focus. While actual breaks in specific parts of the bone do not get passed down to the offspring, there are some features driven by damage that do get passed down to the offspring’s body.
Defining some terms:
• Synostosis is the process where the bones are pushed down into a series of phases, leading to two or more bones thickening from the stress they receive, then merging. The phases of synostosis end when the girthier bones begin overlapping and growing into each other’s spaces once they make contact, climaxing with all of them eventually forming a single solid bone that merges with the connecting bones.
• Homeostasis is when the body is in a state of being in its strongest and most stable condition, rested and repaired, with level biochemistry and optimal immune responses. Returning to homeostasis is a process of damage and repair, as entropy is counteracted by the body’s patch-and-fix response to all pain and injury. The living body is always in a state of repair, even when it is stable. Cellular repair and regeneration are other hidden forces working with evolution. It is the hidden force responsible for providing maintenance against the degenerative force of entropy. Your body’s healing responses patch up the deterioration that biological erosion would otherwise reduce to total destruction of the body and its systems if it weren’t counteracted by repair. The return to homeostasis is what prevents evolution from becoming a chaotic ball of dysfunction. It creates gains that get preserved in the DNA upon reproduction. This drives the evolution of the organism into a never-ending line of fitness, which natural selection also acts upon. When this occurs and is preserved, the potential for advancement becomes exponential—the only thing that stops a species from becoming its best model for its conquered niche is extinction itself, from a surprise shift in the environment or from being outcompeted by a better organism. As entropy increases over time, it triggers life to repair against it and make constant corrections that the body automatically responds to—and this dance between entropy and ectropy increases the potential for advanced lifeforms to emerge out of this natural system.
Both of these biological processes, which can occur in an individual’s lifetime during healing, are processes I have observed being passed down genetically. As we’ve already defined, we are coming to realize now that biological erosion (the strange biophysical concept closely tied to entropy) is a force of gradual wear and tear on anatomical structures due to repetitive behaviors, which lead to injury or degenerative impacts on the body. It would be appropriate to say that this deteriorating force is what drives these two healing forces to respond the way they do, thickening and exaggerating the shape of a bone or tissue. In fact, we already know that there is a connecting relationship between damage and the body’s repair response; this part is not even a theory but an observable fact in your current living body whenever you get injured and watch the wound heal. What is surprising is that this relationship between the two forces continues to shape the bodies of the great-grandchildren of a collective breeding population, all participating in the same repetitive, damaging activities that leave a trail of their activity in the worn-down shape and thickness of an entire lineage’s bones.
This is profound because when you know what to look for, you can actually see how the body builds itself and the exact moment a new behavior kicked in, which is pressed into these shapes by its own movement through life but is also influenced by the environmental pressures to survive.
The external environment causes the animal to move its body only in ways that ensure its survival with the pressures of a particular environment, but if you were to control your environment, or control your behavior, then evolution isn’t a prison but a thing you can directly guide and assist with intentional design behind your direction. So, at the cost of facing your pains for gains and conquering your fears, you can shape your species into anything you want it to become using this mechanism.
Examples of the predictive power of these mechanics:
This process can lead to significant changes in skeletal morphology over time. An animal that is not designed to use its body in a strange but useful way to survive will be met with damage to the imperfect areas of the body that are not yet developed or adapted enough to handle the damage better. For example, let’s look closer at the evolution of the horse’s hoof, where five metacarpal bones and five fingers fused into one single metacarpal and phalanx. This happened to them because the earliest ancestors of the horse were being chased by giant terror birds 38 million years ago through the swampy marshlands of Eurasia. The wet ground made for plenty of stuck hyracotherium and eohippus individuals struggling to run through the mud while getting a leg sucked into the muck. This ultimately made each failure an easy meal for the terror bird that was chasing it into the mud. The only early horses that were able to survive and copy their genes into their young were the ones that could run through the muddy terrain on the tips of their toes to keep the surface area of their foot narrower so they could run faster through the wetlands and avoid getting their five hoofed toes literally stuck in the steps and phases that the family tree underwent.
Hidden Force 4 —Repetitive Movements / Zipf’s Law
Zipf’s Law describes a pattern in which highly popular choices or items dominate in frequency or usage, while the contrary is much less common or rarely chosen. Over countless generations, the accumulation of the horse’s popular choices to run on the tips of their toes over soft ground made the five separate metacarpal and phalangeal bones fuse into a single, stronger hoof, as osteoblasts deposited new bone tissue in response to the continuous stress they were always under. This is a striking example of synostosis being recorded into the genes as gravity caused their middle toe bones to constantly be crushed and healed with thicker osteoblasts, and these life-acquired girth growths of each bone in their paws got thicker as the animal’s entire mass grew in size and weight over the next 35 million years. With each fossil discovered of the horse’s ancestors, it has recorded this slow process of synostosis going through all the stages over the extension of 35 million years; this can only mean it was always getting passed down into the future offspring, which, after being born, only continued to further warp the toes by repeating the same foot posture as their parents maintained; making the two middle toes smashed thicker as the hoof was formed from every stamping foot of the species working as one stampeding unit toward these maximized evolutionary changes. The fact that the two middle digits on each foot fused into one single strong toe bone that was durable enough to support the weight of 1500 lbs. of mass on only four toes together exemplifies biological erosion’s impact when toes are smashed together and flattened at the edges of the hoof for both speed and stability. We can look at the evolution of the horse’s hoof as a way of understanding the predictive power of this new evolutionary theory, which can forecast the future design of a genome through repeated mechanical stimuli observed in the present itself, illustrating a form of acquired trait inheritance driven by movement, injuries, and environmental interaction.

Figure 1: Illustrates the process of synostosis/atrophy in the evolution of the horse’s hoof. Image by Chris Jennings
III. Gravity’s Influence: A Biophysical Perspective
Gravity is one of the most pervasive, yet camouflaged forces hidden behind the scenes, influencing biological evolution. Gravity’s role extends beyond mere support; it actively molds skeletal and muscular systems through predictable, mechanical feedback. The physical forces exerted by gravity induce specific structural adaptations that are often preserved as morphological signatures, such as being crushed, thickened, bent, or all of the above, due to the body’s weight and its orientation relative to the direction of gravity.
From the earliest tetrapods to complex mammals, from walking on all fours or upright, gravitational forces have dictated structural adaptations in every skeletal arrangement, shaping them as necessary for survival, while at the same time being observably warped under a gravity-dominated environment. The transition from aquatic to terrestrial environments posed significant challenges for living organisms used to floating through their environment. This included the need to adapt to gravitational forces on top of breathing. Gravity not only built the first legs for land animals by triggering the bone’s healing properties to remain activated while bending their shapes, but it is a fundamental force of nature that played a profound role in shaping the planet into a sphere, its orbit around the sun, and ultimately the evolution of living organisms; all of which are forced to adapt to every external force exerted on them.
The effects of gravity on skeletal structure are evident in various species. Two of many such great examples where gravity is caught making its mark on the shapes of bones are found astonishingly in humans and turtles.
Humans
The Relationship between Gravity and the Human Spine: The human spine is crunched downward into an S shape. This is no coincidence; it actually looks smashed like a soda can by gravity. No other animal in the animal kingdom has this type of spine; the closest to it are the great apes, but even they don’t have a perfect S-shaped spine built to support a vertical posture better than humans. The S shape looks like a deformity caused by compression downward, and it certainly could have started out that way, but it’s ideally not painful and is comfortably adapted to withstand the compressive forces of gravity while in an upright, vertical gait without abuse. So, we see a pattern where useful shapes that pass through natural selection’s approval are given the power to proceed; it’s like taking snapshots through time, where copies of the whole process the bones went through as they healed, while being bent or worn, are made
The development of each curved shape in our spines distributes our weight away from it; this, in turn, shifts the body’s weight centre, keeping it from placing too much pressure on a single focal point in the spine. It is also flexible, like a spring, and allows better shock absorption when running or jumping, and landing on two legs more efficiently than other animals’ spines, which are built for four legs. In humans, spinal curvature (specifically, lumbar lordosis) is a direct response to the need to balance and distribute the compressive forces exerted by gravity during upright posture. This curvature reduces stress on the vertebral column and minimizes injury over a lifetime of bipedal locomotion.
Not only was the spine shaped by gravity’s continual presence, but human femur bones and pelvises are uniquely shaped in such a way that reveals how standing upright more often through life can thicken the lumbar discs and flatten the ilium area of the pelvis, and bend the femurs inward into a V shape. Again, like the spine, although it also looks like an uncomfortable deformity from being crushed by gravity, the V shape where our knees meet helps us keep our center of weight down the middle of the body to our ankles, instead of wobbling far left and right like a chimp does when it tries to walk upright. These modifications helped us, but the two-legged walking advantage came at a cost: it does make human birth labor way more painful compared to what other placental mammals experience. The V shape of our pelvis compresses the pubic bone, and then add to that the larger head of a human infant trying to squeeze out of it, which can make the birthing process problematic. Compared to all quadrupedal animals, humans seem to have distorted their skeletons from the damage and repairs caused by forcing themselves to walk upright vertically against gravity instead of hunched over horizontally on all fours. They did this when these shapes in our bones were not yet fully developed, and as a result, they were filled with more error and discomfort than what we’ve achieved today. This earlier ancestor of ours, with a more awkward ape-like spine and pelvis, would’ve been alive somewhere around 10–15 million years ago.
Additional notes on Human Evolution:
The traces for bipedalism in early hominins are preserved in the fossilized pelvises and spinal columns of Ardipithecus ramidus, Australopithecines, and surviving archaic Homo species such as Homo naledi. These show the step-by-step adaptations to upright posture, including lumbar lordosis and changes in foot and pelvic structures. Repeated weight-bearing and balancing activities reinforced these features; perhaps bipedality evolved in the trees for us first before it was mastered on the ground. This idea is supported by the anatomical comparisons between Orrorin tugenensis, Ardipithecus ramidus, and the greater Australopithecines closest to the genus homo.
The human foot was 100% shaped by the smudging and warping of its original ape-foot shape simply from walking on two legs rather than using all four limbs together. The weight of the full body was suddenly shifted to bearing all the weight on two legs instead of climbing in the branches with all four limbs or crawling, and our feet took all the damage. Think of their shape like a deformity caused by the walking action itself; if our feet were made of soft clay, the current human shape would have formed in only one lifetime of walking on them (moving from a gorilla-style foot towards a human-shaped foot). Gravity did all of this to us through a process of deterioration mixed with repair on hard bone over millions of years, in the same way that gravity flattens fossils over time under hundreds of tons of layers of rock above them in the Earth’s crust.
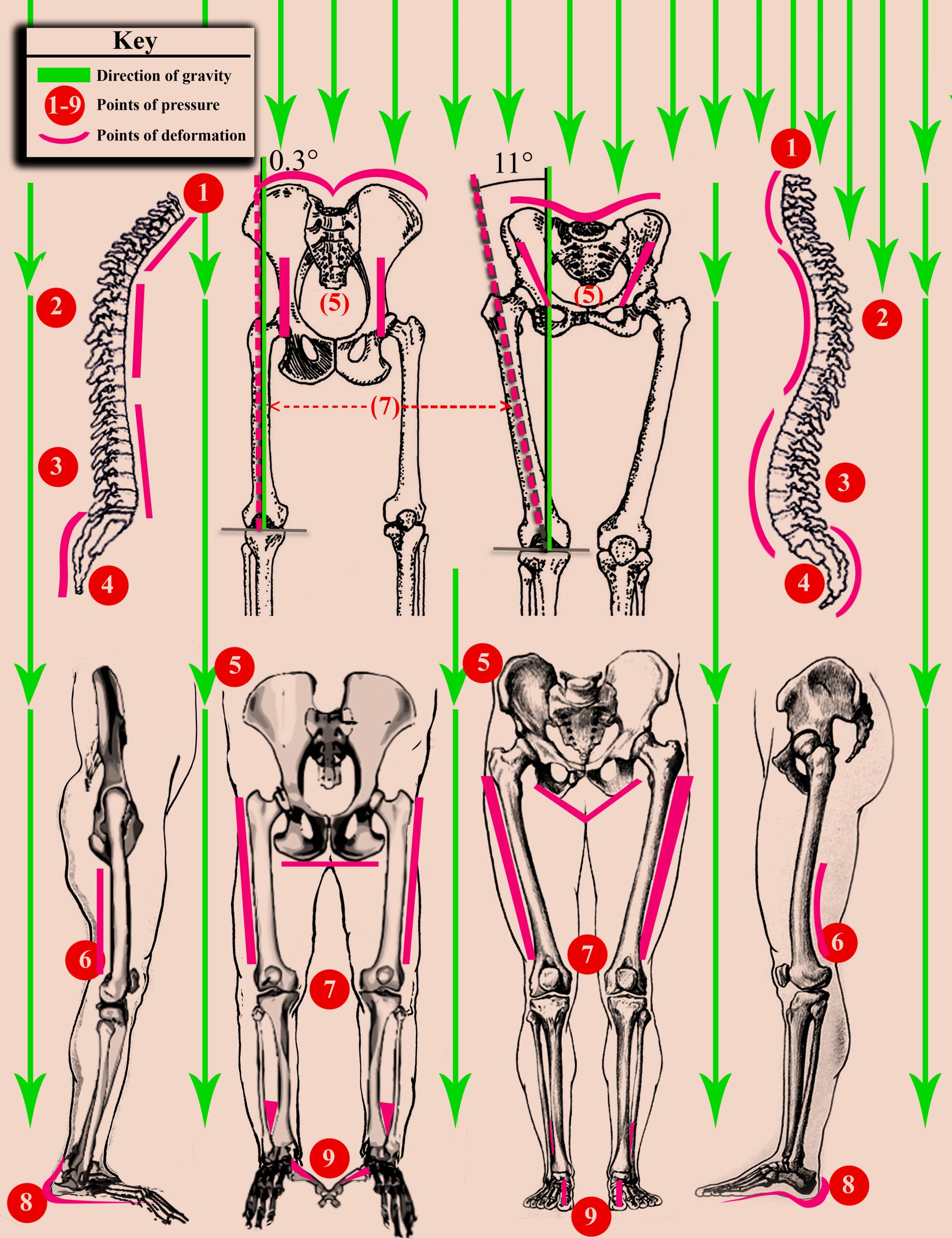
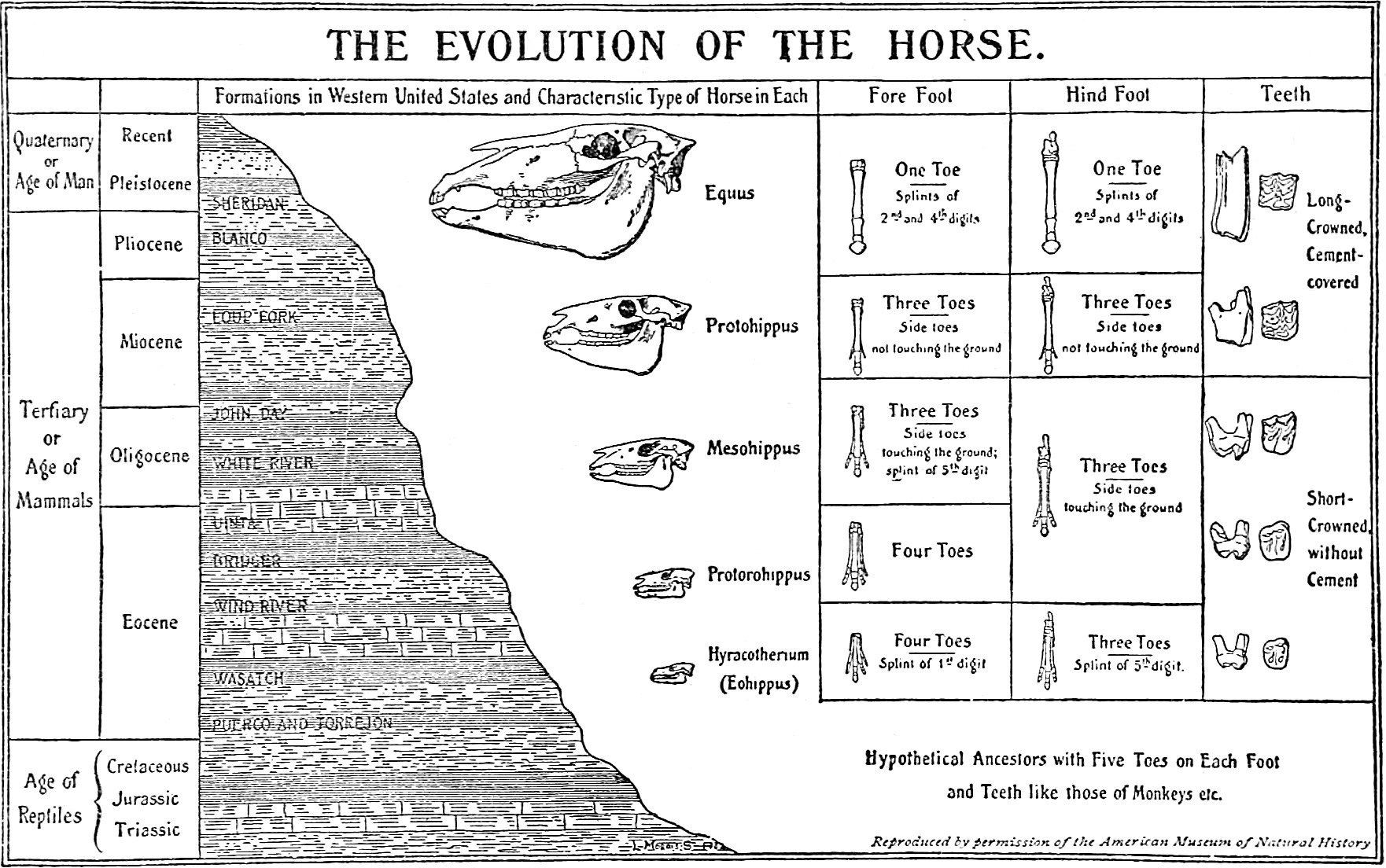
Figure 2: This list identifies specific regions of the human skeleton where gravitational stress influenced anatomical remodeling during our evolutionary transition to bipedalism. These regions exhibit predictable bending and warping consistent with the forces we just observed in early tetrapod humeri. Image by Chris Jennings
Breaking down the meaning of Figure 2
We are comparing the ancestral ape-like morphology (left) to the modern human (right) and the morphological differences that reveal the following (gravity-related) modifications:
(1) Cervical Vertebrae: The cervical spine developed a lordotic curve to support the cranium’s weight in an upright posture. In Figure 2, this can be seen illustrated as the pink curved line under the color-coded (1) located on the right-hand side (human spine). Compare this curve to the (1) on the left-hand side (ape spine) and view it as a before-and-after picture, glancing from the left diagram to the right diagram. This lordotic curve was caused by the constant weight of the head that was pressing down into the base of the neck while human ancestors stood in an upright walking gait, as gravity acted on each cervical disc.
(2) Thoracic Column: The thoracic spine transitioned to a kyphotic curve due to vertical axial loading. Once again, the kyphotic curve relates directly to the weight of both the head and the shoulders, crushing down into the middle back. Our entire spine was crunched downward vertically as a soda pop can under gravity; each curve counterbalanced the one above it and shifted the centre of gravity away from straining one targeted area of the spinal column. This is what makes our spines so well adapted for bipedality; other members of the Great Apes do not have this many curves in their spines, as you can see with the left-hand side comparison in Figure 2.
(3) Lumbar Column: Lumbar vertebrae became more robust and developed a secondary lordotic curve to sustain the weight of the upper torso during bipedal gait. This is the base of the spine before it connects to the pelvis. All the weight of the entire torso is being held up by our lower backs, and the lordotic curve we see in the lumbar column is paired with the thickest vertebral discs because the crushing forces of all the body mass above it has forced the bones to keep healing more frequently at the bottom of the spine during the entire period of time our ancestors were practicing bipedal locomotion. Compare each spinal section in humans with the ape spine on the left, and you see a predictable pattern in which the crushing force of gravity is shaping the exact consequential shapes we would expect to see occur as it sculpts their curves and bends into the bones of the future offspring.
(4) Sacrum and Coccyx: The sacral discs underwent synostosis (fusion), while the coccyx curved anteriorly, likely influenced by pelvic pressure during bipedal walking and the development of robust gluteal muscles. Once again, we see synostosis here, where the pelvis meets the lumbar column, the discs fused into a solid pillar structure we call the sacrum. This occurred because there is no flexibility needed as the pelvis encases these discs in the spine into a rigid structure that holds the integrity of our weight and balance. Constant weight pressure at the base of our spine sent the discs into constant thickening, and wherever flexibility is not needed, you will see synostosis take over and fuse bones together to form a sturdier anchor for the body’s weight to rest on. In this section, we also see that the coccyx (tailbone) has also fused together as it atrophied. The curve we see in the coccyx is also gravity-related, as this is the part of our spines that human ancestors sat down on and pushed inward each time they sat on a rock or on the ground. Although the tailbone is a vestige of an external tail, this bone now serves as the anchoring point for the pelvic floor, extending into the gluteus maximus muscles and sphincter muscles in the anus.
(5) Pelvis: The iliac blades widened and flattened under the weight of the viscera and torso in a vertical position. Constant pressure on the femoral heads and pelvic girdle remodeled the ischium into a narrow, V-shaped configuration. Here we can see that a form of biological erosion has occurred on our ilium, which is the upper wings of the pelvis. When apes walk on 4 legs, the weight of their digestive tracts and torsos is not resting on top of their pelvis; they are resting on soft tissue instead, solely supported by their abdominal muscles. In modern humans and archaic human ancestors, the digestive tract and the entire upper body rest right on the pelvis. When we switched from a horizontal gait to a vertical gait, it made all the difference; now the weight of almost half of our bodies rested right on the pelvis. Every jump, run, and twist of the body eroded the inner walls of the ilium to curve and be more bowl-shaped, which was favored by natural selection because it cradles the intestines much better for a comfortable fit. Then we see the upper wings of the pelvis are flattened down into a squattier shape as compared to the tall and broad wings of a gorilla or chimpanzee’s pelvis. The inner bowl-shaped arrangement of the modern human pelvis is a product of biological erosion as the weight and movement of the inner organs pushed their way downward into the cavity of the pelvis and shaped the bones to be more concave than the more straight up and down iliac sections in an ape’s pelvis.
(6) Distal Femur: The femoral condyles at the knees exhibit an increased curvature from the lateral view compared to non-human primates. From the side view, our thigh bones are curved like the letter C when compared to an ape’s thigh bones. This is, yet again, another morphological feature that got crushed under the force of gravity from just standing upright more often during our active hours of the day. Apes are not standing on two legs for as many hours as our early ancestors did, so they are not receiving any of these severe curvatures in their legs and spines.
(7) Femoral Angle: A distinct 11° bicondylar (valgus) angle developed as the femur remodeled in response to the sinking pelvic girdle, allowing the knees to touch when the feet are together. What we are seeing here is another predictable pattern where gravity’s presence has been recorded into the slow shaping of our evolving bones. When apes are forced to walk on their hind legs, they waddle with wide steps, and this places severe mechanical stress on the knees and the ball-and-socket joints of the femurs. Looking at our legs from the front or back view, we see that our legs are positioned perfectly by natural selection to keep our steps closer together at the center of our balance with gravity. This bowed shape is exactly what we would expect to happen to both the knees and the femoral heads because they’ve constantly been taking a beating from gravity as each leg was forced to carry 100% of the body weight instead of sharing it with 4 legs working together. Humans now have legs that are bowed inward, just the shape of the legs on a weak table would be if you stacked a pile of a thousand tons of bricks on top of it and left it there for years, crushing the 4 legs into a bowed shape.
(8) Pes (Foot) Meta Tarsal Arch and Heel: High-impact activities like sprinting and leaping or tiptoeing on the balls of the toes to stealth sneak on unsuspecting prey created the longitudinal arch (unlike the flat metatarsals of Great Apes) that the human foot has today. The modern human foot also features a robust, rounded calcaneus (heel), unlike the nimble heels of apes. Our heels were rounded out and thickened from constantly walking and running hard on them, with all of our weight bearing on them. Notice how much larger our heels are when they are compared to the significantly smaller acute heels of all the Great Apes.
(9) Lower Leg and Hallux: The tibia and fibula became less flexible for climbing and more rigidly anchored for stability in bipedal locomotion. The hallux (big toe) realigned in an adducted, forward-facing position to serve as a durable lever for propulsion, which appears to have been smudged and smashed into this position by the constant walking habit itself, as gravity worked on every bone along the way. Our big toes were pushed upward into the top of the foot just from the motion of walking alone. The human foot took the most beating from gravity out of every other bone in our body because it is the base of our locomotion and was forced to support 100% of the weight of our entire bodies.
• Bone Density Variations in Astronauts:
It is also important to observe what happens to bones when gravity is reduced or removed. Modern studies of astronauts in microgravity environments reveal rapid bone loss (atrophy) and remodeling; the exact opposite of what occurs when bones are crushed under gravity on the planet’s surface. Instead of becoming stronger and bending or warping under a crushing force, the bones in outer space begin to lose internal strength and become less dense, looser, and more flexible around gaping joints, moving out of place from their normal original positions while living under gravity.
This opposite effect is a good way to troubleshoot the mechanics behind this pattern in evolution; flipping the force on and off like a switch to watch what it does to the body and improve our understanding of how it works in forward or reverse. This also sheds light on why whales lost their limbs underwater. Would humans evolve to lose their limbs too if we adapted to become an outer-space-dwelling creature or a marine-adapted one? This helps illustrate how physical forces (or lack thereof) directly influence bone structure, paralleling how habitual gravity-related stresses shape bones across evolution.
This development of biomechanical adaptation further exemplifies how physical forces, such as gravity, can reshape skeletal morphology and show up in the form of biological erosion and distortion of the body’s shape and layout. These slow-moving processes leave behind a long trail of their progress, and they will always be caught in the act when you compare the changes that have occurred over deep time. They are sneaky to spot, but once you know what to look for, you’ll start seeing them everywhere in the body of any organism.
Turtles
The Relationship between Gravity and the Turtle’s Shell: The turtle’s shell is a merging of the spinal discs, ribs, and the outer skin, forming into a large bone-plated shell that the animal can use as armor to protect itself. The dorsal shell of land tortoises exhibits thickening and reinforcement (synostosis of the ribs and spine) in the areas subjected to habitual rough contact or hard pressure accumulated from either being attacked and surviving, stumbling down cliffs and surviving, burrowing, and mounting friction and movement during their mating behaviors.
The key area of interest is the underside of the shell, known as the plastron. In land tortoises, especially large species such as the Galápagos giant tortoise (Chelonoidis nigra), the shape and robustness of the plastron vary significantly across habitats and under different gravitational influences. Male tortoises develop a concave plastron, which facilitates mounting during mating; a behavior that exerts localized pressure, encouraging bone and shell remodeling in response to mechanical stress. Conversely, females tend to have flatter plastrons, optimized for stability and weight distribution. These morphological differences are not purely inherited; they are reinforced through repeated physical interactions with gravity and environmental forces, leading to structural modifications that are perpetuated across generations.
Figure 3: Shows the underside plastron of a male and female box turtle and further emphasizes the role that repetitive behaviors play in phenotype expression. Image by Chris Jennings
https://www.centralmsturtlerescue.com/uploads/2/3/5/7/23579920/9233587_orig.jpg
{kind=link}
Comparing the male plastron of a sea turtle that mates underwater to that of a giant Galápagos tortoise that mates on land provides an interesting perspective on how gravity (or even the absence of gravity) influences morphological adaptations.
• Sea Turtle (Underwater Mating):
Male sea turtles typically have a moderately concave plastron. This concavity allows them to grip the female securely during underwater copulation, where buoyancy and water resistance create different mechanical demands than on land. The depth of the concavity in sea turtles tends to be relatively shallow (sufficient for gripping but not overly pronounced) because the primary mechanical challenge is maintaining stability in a buoyant, water-based environment. The gravitational forces are less directly felt because water supports much of the turtle’s body weight, reducing the need for deep structural adaptations solely for gravity’s influence.
• Galápagos Tortoise (Land Mating):
In contrast, male Galápagos tortoises exhibit a much more pronounced concave plastron—sometimes significantly deeper—because they mate on land, where gravity exerts a constant, unrelenting force. The deeper concavity functions as a mechanical advantage, allowing the male to mount and maintain position on the female against the pull of gravity. This morphological feature is a more severe adaptation, reflecting the stronger influence of gravity on land. The greater depth of the concavity indicates a higher degree of structural reinforcement necessary to counteract gravitational forces during mating.

Figure 4: A side-by-side comparison of two very large and small male sea turtles’ plastrons versus a male giant Galápagos tortoise’s plastron (measuring the severity of gravity’s impression left on bones). Here we are using the anti-gravity effect that water has on lightening loads to troubleshoot gravity’s role in shaping biology/genetics. If gravity had zero effect in this manner, then the 2,000-lb leatherback sea turtle should have a much more indented plastron than the 600-lb tortoise, but since leatherbacks do not mate on shore in the gravity-influenced environment on land, their plastron is only indented as much as it needs to be for keeping physical contact with their female’s carapace. Therefore, the giant tortoise proves that its weight (on land) makes all the difference, and it has the most indented plastron among all land turtles and tortoises. Image by Chris Jennings
Comparison and Interpretation:
Therefore, yes, the difference in the degree of concavity, the depth of the depression, is more pronounced in the land-mating Galápagos tortoise than in the underwater-mating sea turtle. This suggests that the impact of gravity on morphological features is more severe and demands more substantial structural adaptation in terrestrial environments. The tortoise’s deeper, more concave plastron exemplifies a response to the persistent gravitational challenge, reinforcing the argument that physical forces such as gravity actively shape biological form through functional modifications. One can only conclude that males developed their concave plastrons from the repetitive act of pressing hard into the female’s back against their bellies. Natural selection permitted this dimple to form so that future land turtles could balance on the female’s back better while moving and facilitating a successful mounting during mating, a behavior that exerts localized pressure from gravity and influences the male plastron’s shape. The structural modifications in these shells are not purely genetic but are reinforced and perpetuated by mechanical stresses encountered during life, through repetitive actions related to survival, which then influence bone and shell growth patterns. The different morphologies of male tortoise and sea turtle shells exemplify how physical forces, especially gravity, indeed have a dominant influence on the shape of biological structures beyond only genetic predispositions rooted in genetic determinism.
In Figures 1-4, we are presented with illustrated samples of the many forms of evidence of gravitational influences in evolution working with repetitive motions. In every single case where we would expect to see warping and bending of a weight-based pressure bearing down on weaker bones, we see the correlating curvatures and thickening of structures from those weighted pressures. This then either triggers synostosis to occur or just the thickening of the targeted bone. What breaks it away from the path of fusion is a demand for flexibility (a good example is in joints and wrist bones). Wherever there is no need for flexibility, fusion (synostosis) will occur, and the merging of the bones will be inevitable as long as heavier weight keeps pressing down on the fusion points.
IV. Rethinking Lamarckian Evolution
Hidden Force 5 —Atrophy / Lamarck’s Use & Disuse Model
The last and final hidden force is atrophy/repetitive disuse. When a specific organ is neglected with no frequent use for millions of generations (i.e., like the reduced arms of Tyrannosaurus rex, the hind legs of whales, or the tailbone in humans), the exact opposite effect of synostosis occurs—rather than growing, it shrinks, weakens, and becomes non-functional until it turns into a vestigial organ and, eventually, disappears from the body. This is not very different from what Jean-Baptiste Lamarck originally stated; it is just more refined with up-to-date information about how biological processes occur.
Lamarck’s original concept of his “use and disuse” model was not that far from the truth, although it was worded prematurely in its initial description and understanding. In 1808, Lamarck made his best attempt to describe evolution as a dynamic interplay between an organism’s environment, behavioral choices, and physiological adaptations resulting from those choices. He introduced the idea of a substance he called “nervous fluid” flowing in or out of an organ based on its use or disuse. While this aspect of his theory has been proven false—there is no special fluid causing these changes—his notion of a “nervous fluid” (also called “subtle fluid”) guiding organ growth or atrophy hints at underlying principles that remain relevant.
This idea subtly foreshadows modern concepts like vestigial organs—structures that appear to be shrinking or losing function over time due to infrequent use or diminished need, resulting in morphological changes driven by positive or negative influences. Reinterpreting Lamarck’s old idea through a modern lens highlights concepts such as synostosis and homeostasis, replacing his outdated notion of a flowing “nervous fluid” with the understanding that cellular and physiological processes are the true driving forces behind adaptive growth.
We now know that repetitive use and damage are interconnected; where damage occurs, regeneration and nutrient delivery facilitate repair. Conversely, atrophy—the gradual decline of unused organs—is essentially the reverse of this process, representing neglect to nourish or repair a targeted organ due to prolonged inactivity and stagnation. Therefore, we can move away from Lamarck’s antiquated language of “fluids” flowing and bridge his original ideas to our current understanding rooted in cellular biology and observable physiological processes. This reveals that Lamarck’s core premise—that use and disuse influence organismal form in a family tree—was not entirely incorrect. Instead, it was simply ahead of its time, lacking the terminology and mechanistic insight provided by modern-day biology.
In my third and final article under this same title thread, I will explore which other ideas from Lamarck were correct. I will demonstrate the details of my ceiling fan and feather experiments, which I briefly explained in my first article, and showcase other repeatable experiments I performed to better understand these hidden forces. Part 3 will distill the study into three basic laws that govern the entire system. Once we bring this to a close, it will help humanity understand what causes evolution to occur with greater predictability and confidence—shedding light on what makes our bodies change from what they were, into what they are now, and what they will become in the future.
Thank you for taking the time to learn more about this work! Be sure to catch up on the introduction, and look out for Part 3 of this article thread coming soon!
Bibliography:
(1) West-Eberhard, M. J. (2003). Developmental Plasticity and Evolution. Oxford University Press.
(2) Emerson, S. B., & Koehl, M. A. R. (1990). “The interaction of behavioral and morphological change in the evolution of a novel locomotor type: ‘flying’ frogs.” Evolution, 44(8), 1931–1946.
(3) Lieberman, D. E. (2011). The Evolution of the Human Head. Harvard University Press.
(4) Richmond, B. G., & Jungers, W. L. (2008). “Evolution of the human coxal bone and pelvis.” Anthropological Science, 116(2), 131–146.
(5) Latimer, B. (1996). “The human spine: a review of its evolution, structure, and function.” Journal of Anatomy, 188(2), 255–266.
(6) Prothero, D. R., & Schoch, R. M. (2002). Horns, Tusk, and Flippers: The Evolution of Hoofed Mammals. Johns Hopkins University Press.
(7) National Geographic Society. (1986). The Incredible Machine. Washington, D.C.: National Geographic Society. (Contributors include Lennart Nilsson and Robert M. Poole).
(8) McMinn, R. M. H., Hutchings, R. T., & Abrahams, P. H. (2002). McMinn’s Color Atlas of Human Anatomy (5th ed.). St. Louis, MO: Mosby. (Also known as McMinn’s Clinical Atlas of Human Anatomy).
(9) McMillan, B. (2008). The Illustrated Atlas of the Human Body. Sydney, Australia: Weldon Owen Publishing..
Fossil Record and Specific Taxa mentioned:
Daeschler, E. B., Shubin, N. H., & Jenkins, F. A. (2006). “A Devonian tetrapod-like fish and the evolution of the tetrapod body plan.” Nature, 440(7085), 757–763.
Acanthostega
Clack, J. A. (2012). Gaining Ground: The Origin and Evolution of Tetrapods. Indiana University Press.
Hyracotherium / Eohippus
MacFadden, B. J. (1992). Fossil Horses: Systematics, Paleobiology, and Evolution of the Family Equidae. Cambridge University Press.
Megatherium
McDonald, H. G. (1991). The Giant Ground Sloths. Smithsonian Institution Press.
Thylacosmilus
Wroe, S., et al. (2004). “The Type Species of Thylacosmilus atrox (Metatheria, Thylacosmilidae), and the Evolution of Saber-Tooth Morphology.” Journal of Vertebrate Paleontology, 24(3), 640–654.
Procoptodon
Henry, M., et al. (2014). “The Evolutionary History and Ecology of the Giant Short-Faced Kangaroo Procoptodon.” Paleobiology, 40(3), 451–469.
Ardipithecus ramidus
White, T. D., & Suwa, G. (2010). The Evolution of the Human Hand. Harvard University Press.
Australopithecus
Johanson, D. C., & Edey, M. (1981). Lucy: The Beginnings of Humankind. Simon & Schuster.
Homo naledi
Berger, L. R., et al. (2015). “Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa.” eLife, 4, e09560.
Orrorin tugenensis
Senut, B., et al. (2001). “First hominid from the Miocene (Orrorin tugenensis, Tugen Hills, Kenya).” Comptes Rendus de l’Académie des Sciences – Series IIA – Earth and Planetary Science, 332(2), 135–144.